ABO式血液型

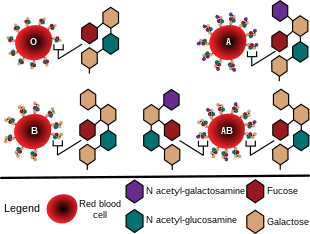

ABO式血液型(ABOしきけつえきがた)とは、血液型の分類法の一種。ヒトの場合はA、B、O、ABの4型に分類する。型を決定する対立遺伝子はA、B、Oの3種、遺伝子型はAA、BB、AB、AO、BO、OOの6種がある。
目次
1 概要
2 歴史
3 機構
4 分布
4.1 地理的分布
4.2 世界の血液型分布
4.2.1 国別ABO式血液型割合
4.2.2 国別ABO・Rh式血液型割合
4.2.3 民族別ABO式血液型割合
4.3 血液型と人類学の歴史
5 ABO式血液型の亜型分類
5.1 A型の亜型
5.2 B型の亜型
5.3 AB型の亜型
5.4 O型の亜型(便宜上ボンベイ型・パラボンベイ型も解説)
6 判定方法
7 抗原側検査
8 血清側検査
9 解離試験・吸収試験
9.1 熱解離
9.2 DT解離
9.3 ジキトニン解離
9.4 グリシン塩酸/EDTA解離
9.5 クロロキン解離法
9.6 ZZAP処理
9.7 PEG吸収
10 汎血球凝集の各レクチンに対する反応
11 ABO型における親子の理論的な血液型の組み合わせ
12 ABO式血液型と性格
13 ABO式血液型と体質
14 ABO式血液型の変化
14.1 後天的な変化
14.2 輸血用ABO抗原の変更
15 脚注
16 出典
17 関連項目
18 外部リンク
概要
血液型は初め血液の型として出発したのでこの名があるが、その後の研究において血液のみに関わらず一個人の細胞、臓器、体液にはもちろん、毛髪などの硬組織にも分布する個人を血清学的に識別できる方法であることが分かっている[1]
赤血球の表面には250種以上の表面抗原があるが、A/B型抗原はその代表的な抗原である。
赤血球の表面にA抗原があるとA型、B抗原があるとB型、AとB両方の抗原があるとAB型、両抗原が無いとO型とする[2]。
逆に血漿中には各抗原に反応する抗体があり、通常A型の血漿中には抗B抗体があり、B型の血漿中には抗A抗体があり、AB型の血漿中には抗A抗体も抗B抗体のどちらも無し、O型の血漿には抗A抗体と抗B抗体両方が存在する[2]。
血漿中の抗体を調べることで血液型を判定することを裏試験ともいう[2]。表面抗原に、それぞれ対応する抗体が反応すると赤血球は凝集してしまう。
歴史
最も初期に発見された血液型分類である。
1900年にオーストリア・ハンガリーのウィーン大学で病理学教室の助手をしていたカール・ラントシュタイナー(Karl Landsteiner, 1868年 - 1943年)は、イギリスの病理学者シャタックの「肺炎患者の血球と血清(それぞれ別人)を混ぜていた際に凝集があった」という報告を聞いて、これが正しいか、正しければ肺炎の診断に利用できないかの追試を行った。
そこで自分を含む22人の健康な人の血液を血球と血清に分けて互いに混ぜ合わせて調べた所、以下のような凝集反応の有無が確認され「凝集は健康な人同士でも起こりうる生理的現象で肺炎診断には使えないが、同じ血清を入れた場合でも血球の持ち主によって凝集するときと凝集しない時がある。」という事に気がついた。
(以下の表は要点を抜粋したもので、残り16人分は省略されている)
- +=凝集、-=非凝集。
- 細胞成分=血球、液性成分=血漿。
| 液性成分/細胞成分 | Storck博士 | Pletschknik博士 | Sturli博士 | Erdheim博士 | Zaritsch氏 | ラントシュタイナー[脚注 1] |
|---|---|---|---|---|---|---|
| Storck博士 | - | + | + | + | + | - |
| Pletschknik博士 | - | - | + | + | - | - |
| Sturli博士 | - | + | - | - | + | - |
| Erdheim博士 | - | + | - | - | + | - |
| Zaritsch氏 | - | - | + | + | - | - |
| ラントシュタイナー | - | + | + | + | + | - |
(山本文一郎『ABO血液型が分かる科学』P4 表1-1「ランドスタイナーの赤血球の凝集実験の結果」より)
これらより凝集のする・しないのグループ分けをするとErdheim博士・Sturli博士、Pletschknik博士・Zaritsch氏、Storck博士・ランドスタイナーと分けられ[3]、ランドスタイナーは「人の血液は3つの群に分けられる」として、「A型」「B型」、「C型(このC型は現在のO型である)」と名付け、翌年の1901年11月14日に論文発表した[4]。
さらにその翌年(1902年)にアルフレッド・フォン・デカステロとアドリアノ・シュテュルリによってこの3群のいずれにも入らない第4の型が追加発表された[5]。
こうした血液の型の呼び方について1906年と1909年にポーランドのヤンスキー、アメリカのモスがそれぞれ独自にこの4つの型を番号で第I〜IV群と呼んだが、お互い順番がバラバラ(I・IVがヤンスキーとモスで逆)でそれぞれの分類を使う人同士に混乱が起き、これとはさらに別に1910年にエミール・フォン・デュンゲルンとルドヴィク・ヒルシュフェルトが4種類の血液について詳しい研究をして「人の血液中には凝集原(抗原)AとB、それぞれに反応する抗体αとβがある。」とまとめ、「A抗原を持っている人をA型(抗体はβ)」というようにまとめ、両方の抗原を持つ人をAB型、両方の抗原がない人をO型と呼んだ[6]。
(なお、C型をO型に変更した理由は、数字の「0」(ゼロ)ではなく、ドイツ語の「ohne」(「 - ない」などの意味)の頭文字である可能性が指摘されている[7]。)
その後1928年の国際連盟の血清標準委員会で、フォン・デュンゲルンとヒルシュフェルトの名称を国際的に使うことが決められた[8]。
その後、フォン・デュンゲルンとヒルシュフェルトは1911年に72家族348人の血液型を調べた結果、血液型は遺伝要素があるという仮説を唱えた。
フォン・デュンゲルンおよびヒルシュフェルトによる調査(1911)[9][脚注 2]
| 両親の組み合わせ | 家族数 | Oの子 | Aの子 | Bの子 | ABの子 |
|---|---|---|---|---|---|
| O×O | 11 | 26 | 0 | 0 | 0 |
| A×A | 10 | 11 | 9 | 0 | 0 |
| O×A | 30 | 24 | 57 | 0 | 0 |
| B×B | 1 | 0 | 0 | 2 | 0 |
| O×B | 3 | 3 | 0 | 5 | 0 |
| A×B | 8 | 1 | 9 | 6 | 4 |
| O×AB | 4 | 2 | 2 | 2 | 3 |
| A×AB | 2 | 0 | 3 | 1 | 2 |
| B×AB | 3 | 4 | 2 | 4 | 2 |
| AB×AB | 0 | 0 | 0 | 0 | 0 |
| 計 | 72 | 71 | 102 | 20 | 11 |
これにより「凝集原を持つ人は必ず親がそのタイプの凝集原を持つ」、「両親ともに凝集原のないO型の場合は子供は必ずO型」という法則を知り、これをもとに血液型はメンデルの遺伝法則に従って遺伝して「Aとa」「Bとb」という二対の対立因子を考えた。(二対対立因子説)
二対対立因子説による4血液型の因子型[10]
| 表現型 | 因子型 |
|---|---|
| O型 | aabb |
| A型 | AAbb、Aabb |
| B型 | aaBB、aaBb |
| AB型 | AABB、AABb、AaBB、AaBb |
二対対立因子説による遺伝[11]
| 両親の組み合わせ | 生まれる子供[脚注 3] |
|---|---|
| O×O | O |
| A×A | A、O |
| O×A | A、O |
| B×B | B、O |
| O×B | B、O |
| A×B | A、B、O、AB |
| O×AB | A、B、O、AB |
| A×AB | A、B、O、AB |
| B×AB | A、B、O、AB |
| AB×AB | A、B、O、AB |
一方、1925年にドイツのゲッティンゲン大学の数学者フェリックス・ベルンシュタインは、「二対対立因子説で予測できる数値に比べ現実のAB型が少なすぎる」という理由から、同年日本の古畑種基・市田賢吉・岸孝義(以下、「古畑ら」と表記)は親にAB型がいる場合、二対対立因子説ではいかなる型の子も生まれる(=親子鑑定に使えない)はずだが、調べているうちに逆に「AB型の親からO型の子がいる例が見当たらない[脚注 4]」という理由から別々に二対対立因子説が誤りではないかと推測し、お互いに「三複対立因子説」を提唱した。
(三複対立因子説の詳細については因子の組み合わせは「機構」、遺伝は「ABO型における親子の理論的な血液型の組み合わせ」を参照。)
両者の内容には差異があるが(後述)どちらもO型は「対立因子がない」のではなく「対立因子の1つを持つが他の因子に劣性遺伝する(古畑の言葉を借りると「AとBに対し潜性するa・bが同じもの[12]」)」という考えで、これならばAB型が少ない理由も子供の生まれる組み合わせも説明できるため、確認のため別の学者たちが何度か家族調査を行った結果、日本国内で合計で958家族、3954人を調べ三複対立因子説の通りの調査成績になったため、1927年のアムステルダムで開かれた第3回国際人類学会とベルリンの第5回国際遺伝学会で古畑らはこれを報告し、ついで1933年の第5回太平洋学術会議で1595家族6826人(子供3636人)の調査結果を報告し、血液型の遺伝は三複対立因子説で説明されるようになった[13]。
なお、ベルンシュタインと古畑らの三複対立因子説の一番大きな違いは「O型に型特異性の抗原性があるか否か」についてで、ベルンシュタインはR凝集原とそれに反応する抗R凝集素(ρ)を仮説として唱え、古畑は抗原は合っても全部の型に共通の基本型抗原だとした[脚注 5][14]。
(実際のO型血球に存在したH抗原はAB型血球にも存在するため、ベルンシュタインのいうR凝集原ではなく古畑の言う基本型抗原に近い。)
機構
A型はA型転移酵素をコードする遺伝子を持っており、この酵素が元になるH物質にN-アセチルガラクトミンをつけてA抗原を作るのに対し、B型はB型転移酵素をコードする遺伝子を持っていてこちらの酵素はガラクトースをつけB抗原を作る。AB型は両方の遺伝子を持っているためAとB双方の抗原を作るが、O型はどちらも作れないのでH物質のままになる[15]。
これらの抗原が最初に血液から発見されたために「血液型」という名称を冠するもので、血液以外にも唾液・精液など、すべての体液にも存在する。ただし血球などに抗原をつける遺伝子と唾液などの体液に抗原をつける遺伝子は別系統(後述)なので、1/4の人は後者の遺伝子が働かない非分泌型という血液以外の体液に抗原が出ない(もしくは微量で検出されない)体質である。
ABO式血液型を決定する抗原を作る遺伝子は第9染色体に存在し、通常のA型とB型の遺伝子では両方355個分(厳密には最後が終止コドンなので354個のアミノ酸になる)のアミノ酸のデータでそれぞれA抗原とB抗原の転移酵素を作り、これでH抗原を作った後追加の糖をつけるが、A型とB型では中~後半部に7か所(99・176・219・235・266・268・310番目)作られるアミノ酸が違うものがあり、この違いでA酵素はN-アセチルガラクトミン、B酵素はガラクトースがそれぞれH抗原につけられ、これがA抗原とB抗原の違いになっている。O型の遺伝子の場合は、AやBの遺伝子の後半部分が機能しておらず途中で終止コドンになって、通常のO型では87番目のアミノ酸用の塩基配列が1つ抜けているので以後がずれ、118個分(最後は終止コドンなのでアミノ酸の数は117個)のアミノ酸のデータで酵素を作るため追加の糖が付けられなくなっている[16]。
一方、H物質発現を作る遺伝子は第19染色体に位置し、H前駆物質をH物質へ変換させる。後述のボンベイ型とパラボンベイ型は、H物質を組み立てる際にフコースをつける工程でこの酵素が作られない遺伝子のため「H物質自体が完成せずに、ここから先のA抗原とB抗原も作られない。」というものだが、分泌型か非分泌型かでさらに違いが生じており、非分泌型のボンベイ型ではこれらの抗原が血球以外にも存在しないが、分泌型のパラボンベイ型では体液分泌細胞や上皮細胞用の抗原を作る1型糖鎖(血液細胞や血管壁用の抗原の糖鎖は2型)を作る遺伝子は別系統(分泌・非分泌型の遺伝子)で機能しているため、体液中にはH物質があり、遺伝子によってはAやBの抗原も保有している他、そこから漏れたこれらの抗原が赤血球に吸着されて「わずかだがABOに関する抗原を持つ赤血球」を持つことがある[17]。
- A型 - A遺伝子をすくなくとも一つ持ち、B遺伝子は持たない(AA型、AO型)→A抗原を持つ。B抗原に対する抗体βが形成
- B型 - B遺伝子をすくなくとも一つ持ち、A遺伝子は持たない(BB型、BO型)→B抗原を持つ。A抗原に対する抗体αが形成
- O型 - A遺伝子・B遺伝子ともに無い(OO型)→H抗原のみ持つ。A,B抗原それぞれに対する抗体α、抗体βが形成
- AB型 - A遺伝子・B遺伝子を一つずつ持つ(AB型)→A抗原、B抗原両方を持つ。抗体形成なし
A抗原とB抗原は、持っていないとそれに対する自然抗体が形成されることが多く、この場合、型違い輸血により即時拒絶が起こる。自然抗体がなくとも型違い輸血により1週間程度で新しいIgM抗体が生産されこれが拒絶反応をおこす。そのため、基本的には型違い輸血は行われない。輸血される血液は受血者の血液より少量のため、血漿によって希釈されて抗原抗体反応が起こらなくなる。そのため、かつてはO型は全能供血者、AB型は全能受血者と呼ばれていたが、ABO以外の型物質(Rh因子やMN式血液型など)が存在することもあり現在では緊急時を除いては通常行われない。2010年4月には大阪大学医学部附属病院で治療を受けた60代の患者が同型の赤血球製剤とO型の新鮮凍結血漿の輸血後に死亡する事故が発生している(但し、この患者は搬送当時すでに意識がなかったことから輸血が原因でない可能性もある)[18]。
なお、「自然抗体」というのはRh型のように不適合輸血を受けた後などに抗体を生じる「免疫抗体」に対する呼び方だが、実際にはこれも免疫機構によるもので、1950年代にペンシルバニア大学のゲオルク・スプリンガーの実験で無菌状態で育てたヒヨコはABO式血液型の抗原を持っていない[19]が、B抗原を持つ大腸菌を投与すると抗B抗体を持つようになったという報告があることや、人間も腸内細菌のいない新生児の頃は抗A抗体や抗B抗体をもたないが、生後3~6か月ほどで持つようになることなどから腸内細菌などに対する免疫の結果生じた抗体とされる[20]。
分布
地理的分布
ABO式血液型の遺伝子分布は母集団(地域や人種)によって差が大きく、コロンブス以前の分布[脚注 6]では、O型遺伝子の率は世界的にどこでも多い[脚注 7]が特にアメリカ大陸の先住民[脚注 8]で、中南米ではO型が100%近くになる地域がある。
A型遺伝子が多い地域はヨーロッパ(約25~35%)とオーストラリアの南部と西部の先住民(約30~35%)、また北米でもカナダ・アルバータ州からアメリカ・モンタナ州にかけてのブラッド族・ブラックフィート族(黒足族)と呼ばれる先住民集団[脚注 9]は例外的にA型遺伝子の比率が高い(最大50%以上)。ただ、この3か所はMN式血液型で調べると分布が全く異なる[21]他、後述のB型の分布も大きく異なる。
B型遺伝子が多い地域は北部インドからモンゴル(約30%)となり[脚注 10]、逆に南北アメリカ大陸やオーストラリア先住民では極めて少なく(ほぼ0%)、同じモンゴロイド系の人種でもアジア側とアメリカ側で結果が大きく違うため、B型は「アメリカ先住民の新大陸移住(約2万年~1万5千年前)後にアジアで増加した」か「偶然アメリカ先住民の先祖にB型がほとんどいなかった」のどちらかの可能性が高いと考えられる[22]。
なお、アフリカ大陸のABO型分布には取り立てて特徴がなく、大半の地域で一番多いのはO型(65~75%)だが特にこの地域が多いわけではなく、シベリアやヨーロッパ西沿岸部とさほど変わらず、A型はアジアの大半地域並み(15~25%)・B型は東ヨーロッパ(10~15%)程度である[23]
日本人のABO式血液型の分布はおよそO型32%、A型37%、B型22%、AB型9%だが、古畑種基らの研究によるとわずかに地域差があり、九州・四国・中国ではA型、東北・北陸ではO型がわずかに多く、それぞれ反対の方がその分少ない[脚注 11]という分布の勾配があった。
世界の血液型分布
国別ABO式血液型割合
| 国家 | A型 | B型 | O型 | AB型 |
|---|---|---|---|---|
| 38% | 19% | 35% | 8% | |
| 26.2% | 22.0% | 47.4% | 4.4% | |
| 47% | 8% | 41% | 4% | |
| 26.65% | 23.72% | 43.57% | 6.06% | |
| 45% | 16% | 33% | 6% | |
| 49% | 8% | 39% | 4% | |
| 27.01% | 33.75% | 30.31% | 8.93% | |
| 23.5% | 39.8% | 27.6% | 9.2% | |
| 23% | 25% | 46% | 6% | |
| 37% | 14% | 45% | 4% | |
| 44% | 14% | 36% | 6% | |
| 43% | 11% | 41% | 5% | |
| 44% | 17% | 31% | 8% | |
| 42% | 10% | 45% | 3% | |
| 25% | 21% | 50.5% | 3.5% | |
| 45% | 9% | 43% | 3% | |
| 44% | 11% | 40% | 5% | |
| 45% | 12% | 39% | 4% | |
| 19% | 28% | 41% | 12% | |
| 36% | 17% | 40% | 7% | |
| 30% | 6% | 60% | 2% |
国別ABO・Rh式血液型割合
| 国 | 人口[42] | O+ | A+ | B+ | AB+ | O− | A− | B− | AB− |
|---|---|---|---|---|---|---|---|---|---|
| 21,262,641 | 40.0% | 31.0% | 8.0% | 2.0% | 9.0% | 7.0% | 2.0% | 1.0% | |
| 8,210,281 | 30.0% | 33.0% | 12.0% | 6.0% | 7.0% | 8.0% | 3.0% | 1.0% | |
| 10,414,336 | 38.0% | 34.0% | 8.5% | 4.1% | 7.0% | 6.0% | 1.5% | 0.8% | |
| 198,739,269 | 36.0% | 34.0% | 8.0% | 2.5% | 9.0% | 8.0% | 2.0% | 0.5% | |
| 33,487,208 | 39.0% | 36.0% | 7.6% | 2.5% | 7.0% | 6.0% | 1.4% | 0.5% | |
| 1,339,724,852 | 49.7% | 29.8% | 18.9% | 5.0% | 0.3% | 0.2% | 0.1% | 0.03% | |
| 10,532,770 | 27.0% | 36.0% | 15.0% | 7.0% | 5.0% | 6.0% | 3.0% | 1.0% | |
| 5,500,510 | 35.0% | 37.0% | 8.0% | 4.0% | 6.0% | 7.0% | 2.0% | 1.0% | |
| 1,315,819 | 29.5% | 30.8% | 20.7% | 6.3% | 4.3% | 4.5% | 3.0% | 0.9% | |
| 5,250,275 | 27.0% | 38.0% | 15.0% | 7.0% | 4.0% | 6.0% | 2.0% | 1.0% | |
| 62,150,775 | 36.0% | 37.0% | 9.0% | 3.0% | 6.0% | 7.0% | 1.0% | 1.0% | |
| 82,329,758 | 35.0% | 37.0% | 9.0% | 4.0% | 6.0% | 6.0% | 2.0% | 1.0% | |
| 306,694 | 47.6% | 26.4% | 9.3% | 1.6% | 8.4% | 4.6% | 1.7% | 0.4% | |
| 1,236,344,631 | 35.1% | 21.7% | 30.5% | 7.3% | 2.0% | 1.2% | 1.7% | 0.4% | |
| 4,203,200 | 47.0% | 26.0% | 9.0% | 2.0% | 8.0% | 5.0% | 2.0% | 1.0% | |
| 7,233,701 | 32.0% | 34.0% | 17.0% | 7.0% | 3.0% | 4.0% | 2.0% | 1.0% | |
| 127,368,088 | 29.8% | 37.8% | 21.9% | 9.9% | 0.15% | 0.2% | 0.1% | 0.05% | |
| 73,000,000 | 36.6% | 32.8% | 21.0% | 9.0% | 0.4% | 0.2% | 0.09% | 0.03% | |
| 16,715,999 | 39.5% | 35.0% | 6.7% | 2.5% | 7.5% | 7.0% | 1.3% | 0.5% | |
| 4,213,418 | 38.0% | 32.0% | 9.0% | 3.0% | 9.0% | 6.0% | 2.0% | 1.0% | |
| 4,660,539 | 34.0% | 40.8% | 6.8% | 3.4% | 6.0% | 7.2% | 1.2% | 0.6% | |
| 99,863,000 | 44-46% | 22-23% | 24-25% | 4-6% | <1% | <1% | <1% | <1% | |
| 38,482,919 | 31.0% | 32.0% | 15.0% | 7.0% | 6.0% | 6.0% | 2.0% | 1.0% | |
| 10,707,924 | 36.2% | 39.8% | 6.6% | 2.9% | 6.0% | 6.6% | 1.1% | 0.5% | |
| 28,686,633 | 48.0% | 24.0% | 17.0% | 4.0% | 4.0% | 2.0% | 1.0% | 0.3% | |
| 49,320,000 | 39.0% | 32.0% | 12.0% | 3.0% | 7.0% | 5.0% | 2.0% | 1.0% | |
| 47,125,002 | 36.0% | 34.0% | 8.0% | 2.5% | 9.0% | 8.0% | 2.0% | 0.5% | |
| 9,059,651 | 32.0% | 37.0% | 10.0% | 5.0% | 6.0% | 7.0% | 2.0% | 1.0% | |
| 76,805,524 | 29.8% | 37.8% | 14.2% | 7.2% | 3.9% | 4.7% | 1.6% | 0.8% | |
| 61,113,205 | 37.0% | 35.0% | 8.0% | 3.0% | 7.0% | 7.0% | 2.0% | 1.0% | |
| 307,212,123 | 37.4% | 35.7% | 8.5% | 3.4% | 6.6% | 6.3% | 1.5% | 0.6% | |
| 加重平均 | 2,744,996,114 | 41.9% | 31.2% | 15.4% | 4.8% | 2.9% | 2.7% | 0.8% | 0.3% |
50.0%以上
40.0–49.9%
30.0–39.9%
20.0–29.9%
10.0–19.9%
5.0–9.9%
民族別ABO式血液型割合
| 民族別ABO式血液型割合[72] | ||||
|---|---|---|---|---|
| 民族集団 | O (%) | A (%) | B (%) | AB (%) |
先住民 | 61 | 39 | 0 | 0 |
アビシニア人 | 43 | 27 | 25 | 5 |
アイヌ(日本) | 17 | 32 | 32 | 18 |
アルバニア人 | 38 | 43 | 13 | 6 |
| 大アンダマン人 | 9 | 60 | 23 | 9 |
アラブ人 | 34 | 31 | 29 | 6 |
アルメニア人 | 31 | 50 | 13 | 6 |
アジア系アメリカ人 | 40 | 28 | 27 | 5 |
オーストリア人 | 36 | 44 | 13 | 6 |
バントゥー | 46 | 30 | 19 | 5 |
バスク人 | 51 | 44 | 4 | 1 |
ベルギー人 | 47 | 42 | 8 | 3 |
ボロロ人(ブラジル) | 100 | 0 | 0 | 0 |
ブラジル人 | 47 | 41 | 9 | 3 |
ブルガリア人 | 32 | 44 | 15 | 8 |
ビルマ族 | 36 | 24 | 33 | 7 |
ブリヤート人(シベリア) | 33 | 21 | 38 | 8 |
サン人 | 56 | 34 | 9 | 2 |
中国人-広東人 | 46 | 23 | 25 | 6 |
中国人-北京人 | 29 | 27 | 32 | 13 |
チュヴァシ人 | 30 | 29 | 33 | 7 |
クロアチア人 | 42 | 34 | 17 | 7 |
チェコ人 | 30 | 44 | 18 | 9 |
デンマーク人 | 41 | 44 | 11 | 4 |
オランダ人 | 45 | 43 | 9 | 3 |
エジプト民族 | 33 | 36 | 24 | 8 |
イングランド人 | 47 | 42 | 9 | 3 |
エスキモー (アラスカ) | 38 | 44 | 13 | 5 |
エスキモー (グリーンランド) | 54 | 36 | 23 | 8 |
エストニア人 | 34 | 36 | 23 | 8 |
フィジー人 | 44 | 34 | 17 | 6 |
フィン人 | 34 | 41 | 18 | 7 |
フランス人 | 43 | 47 | 7 | 3 |
グルジア人 | 46 | 37 | 12 | 4 |
ドイツ人 | 41 | 43 | 11 | 5 |
ギリシャ人 | 40 | 42 | 14 | 5 |
ジプシー(ハンガリー) | 29 | 27 | 35 | 10 |
ハワイ先住民 | 37 | 61 | 2 | 1 |
ヒンドゥー教徒 (ボンベイ) | 12 | 29 | 48 | 11 |
マジャル人 | 36 | 43 | 16 | 5 |
アイスランド人 | 56 | 32 | 10 | 3 |
インド人(インド) | 17 | 22 | 53 | 7 |
ネイティブアメリカン | 79 | 16 | 4 | 1 |
アイルランド人 | 52 | 35 | 10 | 3 |
イタリア人(ミラノ) | 46 | 41 | 11 | 3 |
日本人 | 30 | 38 | 22 | 10 |
ユダヤ人(ドイツ) | 42 | 41 | 12 | 5 |
ユダヤ人(ポーランド) | 33 | 41 | 18 | 8 |
カルムイク人 | 26 | 23 | 41 | 11 |
キクユ族(ケニヤ) | 60 | 19 | 20 | 1 |
朝鮮民族 | 28 | 32 | 31 | 10 |
サーミ人 | 29 | 63 | 4 | 4 |
ラトビア人 | 32 | 37 | 24 | 7 |
リトアニア人 | 40 | 34 | 20 | 6 |
マレー人 | 62 | 18 | 20 | 0 |
マオリ | 46 | 54 | 1 | 0 |
マヤ人 | 98 | 1 | 1 | 1 |
モロス | 64 | 16 | 20 | 0 |
ナバホ族 | 73 | 27 | 0 | 0 |
ニコバル人 | 74 | 9 | 15 | 1 |
ノルウェー人 | 39 | 50 | 8 | 4 |
パプア人 (ニューギニア島) | 41 | 27 | 23 | 9 |
ペルシア人 | 38 | 33 | 22 | 7 |
ペルー先住民 | 100 | 0 | 0 | 0 |
フィリピン人 | 45 | 22 | 27 | 6 |
ポーランド人 | 33 | 39 | 20 | 9 |
ポルトガル人 | 35 | 53 | 8 | 4 |
ルーマニア人 | 34 | 41 | 19 | 6 |
ロシア人 | 33 | 36 | 23 | 8 |
サルデーニャ人 | 50 | 26 | 19 | 5 |
スコッツ | 51 | 34 | 12 | 3 |
セルビア系(セルビア人) | 38 | 42 | 16 | 5 |
ションペン人 ニコバル系 | 100 | 0 | 0 | 0 |
スロバキア人 | 42 | 37 | 16 | 5 |
南アフリカ人 | 45 | 40 | 11 | 4 |
スペイン人 | 38 | 47 | 10 | 5 |
スーダン人 | 62 | 16 | 21 | 0 |
スウェーデン人 | 38 | 47 | 10 | 5 |
スイス人 | 40 | 50 | 7 | 3 |
タタール | 28 | 30 | 29 | 13 |
タイ人 | 37 | 22 | 33 | 8 |
テュルク系民族 | 43 | 34 | 18 | 6 |
ウクライナ人 | 37 | 40 | 18 | 6 |
黒人アメリカ人 | 49 | 27 | 20 | 4 |
白人アメリカ人 | 45 | 40 | 11 | 4 |
キン族 | 42 | 22 | 30 | 5 |
平均 | 43.91 | 34.80 | 16.55 | 5.14 |
標準偏差 | 16.87 | 13.80 | 9.97 | 3.41 |
血液型と人類学の歴史
(注:「民族」は本来文化的なもので人種とは無関係だが、この項の『血液型の話』からの引用部位では「民族」を人種の意味で使用している部位がある。)
地域による血液型比率の違い血液型を人類学に最初に応用したのはルードヴィヒ・ヒルシュフェルド夫妻で、1次大戦終結時にマケドニアに集まった各国の兵士8500人の血液型を調べた際、人種の相違によって比率が大きく違う(下図参照)ことを発見し、1918年5月5日にサロニカの医学会で報告。その後いろいろあって遅れたものの1919年10月18日にイギリスの著名な医学雑誌「ランセット誌」に掲載された。
『ヒルシュフェルドらの分類』[73]
| 民族名(人数) | O型% | A型% | B型% | AB型% | 民族示数 |
|---|---|---|---|---|---|
| イギリス人(500) | 46.4 | 43.4 | 7.2 | 3.1 | 4.5 |
| フランス人(500) | 43.2 | 42.6 | 11.2 | 3.0 | 3.2 |
| イタリア人(500) | 47.2 | 38.0 | 11.0 | 3.8 | 2.8 |
| ドイツ人(500) | 40.0 | 43.0 | 12.0 | 5.0 | 2.8 |
| オーストリア人(?) | 42.0 | 40.0 | 10.0 | 8.0 | 2.6 |
| ブルガリア人(500) | 39.0 | 40.6 | 14.2 | 6.2 | 2.6 |
| セルビア人(500) | 38.0 | 41.8 | 15.6 | 4.6 | 2.5 |
| ギリシャ人(500) | 38.2 | 41.6 | 16.2 | 4.0 | 2.5 |
| トルコ人(500) | 36.8 | 38.0 | 18.6 | 6.6 | 1.8 |
| アラビア人(500) | 43.6 | 32.4 | 19.0 | 5.0 | 1.5 |
| ロシア人(500) | 40.7 | 31.2 | 21.8 | 6.3 | 1.3 |
| ユダヤ人(500) | 38.8 | 33.0 | 23.2 | 5.0 | 1.3 |
| マダガスカル人(400) | 43.5 | 26.2 | 23.7 | 4.5 | 1.09 |
| セネガル黒人(500) | 43.2 | 22.6 | 29.0 | 5.0 | 0.8 |
| アンナン人(500) | 42.0 | 22.4 | 28.4 | 7.2 | 0.8 |
| インド人(1000) | 31.0 | 19.0 | 41.0 | 8.5 | 0.56 |
- (注:オーストリア人の人数は全体が8500人なら600人のはずだが、出典の表で「?」とある。)
- 「民族示数」は「A型とAB型の合計百分率を分子」で「B型とAB型の合計百分率を分母」にした数値。A遺伝子を持つ人が多いほど大きくなる。
当時は血液型が個人で不変かどうか自体不明瞭であった(上記の兵士たちは基本的に似たような環境で同じものを食べていたが、もっと長期間の環境変化ではどうなるかは分かっていなかった。)ため、これが彼らの故郷の環境によるものか血統的によるものかがはっきりしなかったため重要視されなかったが、1921年ハンガリー(原書は「ハンガリヤ」表記)のヴェルザーとウェスツェッキーが自国内で当時出自が違う民族とすでに分かっていたドイツ系・蒙古系(注:マジャール人の事)・ジプシー(インド系)の民族の血液型比率を調べたところ、下図のようになった。
『ヴェルザー、ウェスツェッキーの研究』[74]
| 民族名(人数) | O型% | A型% | B型% | AB型% | 民族示数 |
|---|---|---|---|---|---|
| ドイツ系ハンガリア人(476) | 40.8 | 43.5 | 12.6 | 3.1 | 2.9 |
| 蒙古系ハンガリア人(1500) | 31.0 | 38.0 | 18.6 | 12.2 | 1.6 |
| ジプシー(386) | 34.2 | 21.1 | 39.9 | 5.8 | 0.6 |
これらの違いとドイツ系民族やジプシーはヒルシュフェルドのドイツ人やインド人との報告とほぼ同じ比率になったことなどから、離れた国に長期間住んでいても他との通婚が少ない場合血液型の比率が先祖とほぼ変わらないこと、逆に同じ国にいても先祖が違う集団は違ったままであることが判明した[75]。
こうした結果より、民族の移動などを血液型から推測する研究がされるようになり、まず最初のヒルシュフェルド夫妻は『ヒルシュフェルドらの分類』の表のデータのうち、イギリス人からギリシャ人までを「ヨーロッパ型」、マダガスカル人からインド人までを「アフリカ・アジア型」としてその間の4民族を中間型とし、O型の比率はどこも極端に違わないのにA型とB型の人種差が極端に激しいので、「かつてA型を基本とするA人種(ヨーロッパの中~北部起源)とB型を基本とするB人種(インド北方起源)がいて、これが混血していき様々なABO式血液型の比率を生み出した。」という説を提唱した[76]。
その後、世界各地の人種の血液型比率を調べていた際にスナイダーが混血が少ないアメリカ先住民族(原文は「アメリカ・インディアン」)にO型が極めて多い(453人を調べてOが91.3%だった)ことを報告し、これにより自分が提唱した後述の原則から「大昔はアメリカ先住民は100%O型だったのではないか」という説を提唱し[77]、これ以外にドイツのベルンシュタインなども自分の三因子仮説などから原始人類の血液型はO型のみでそこからA・B型が突然変異したのではないかという説を上げていた[78]が、カナダのアルバータ州でマトソンとシュラーデルが調査したところ、ほとんど混血のない先住民(前述のブラッド族・ブラックフィート族など)にA・O型が多くB・AB型が皆無という地域が見つかった他、オーストラリアでも白人との混血が少ないにもかかわらずA型の多い集団が発見され、原始人類はO型のみではないかという仮説は訂正された[79]。
スナイダーは血液型を人類学に応用する際、以下の必要な四原則を定めている。
- 一民族の血液型分布率は、これと血液的に緊密な関係にある他民族の血液型の分布率と近似することが予期される。
- 一民族の血液型分布率が、これと血液的に緊密な関係にある他民族の血液型の分布率と大いに異なる場合、この民族はさらに他の民族との間に混血があることが予期される。
- 一民族の血液型分布率が、これと緊密な関係にない他民族の血液型の分布率と等しい場合、その祖先の時代において前者が後者あるいはその近い民族との間に混血があったことが想像される。
- 一民族がAまたはBのいずれか、あるいは両方を欠くか、その遺伝子頻度が非常に小さい場合、この民族は人類にAまたはBの突然変異が起こる前、またはAやBの広がる前からほかの民族と孤立して生存していたと考えられる。
この四原則は当てはまる場合も多いが、一致しない場合もあり他の証拠から近縁でさらに別の民族との交流が薄い民族同士でも分布率が大きく違い、逆に無関係のはずの2つの民族の血液型分布がほぼ一緒という場合もあるので、なるべく一定数以上の人数で広い範囲を確かな診断で調べる必要がある[80]。
ABO式血液型の亜型分類
ABO型の各型の凝集力の違いなどを元にさらに下の亜型がある。検査については亜型検査を参照。
血液型の亜型はA2が最初の発見になり、通常のA型はフジマメ科の植物ドリコス・ビフローラスからとれるレクチンで凝集が起きる(B型・O型は凝集しない)が、A型であるにもかかわらずこれに反応しないものがあったことで発見された。このA2は酵素反応してないH物質が多く、このためドリコスレクチンに反応しなかった。
原因はA2の遺伝子はABO血液型物質を作る354番目のアミノ酸の塩基配列が1つ抜けたため、次が終止コドンにならずにA1(通常のA)より長くなり、376個分のアミノ酸のデータで酵素を作るためこのような違いが起こっていた。なお後に判明した他の亜型の場合もA3は291番目のアミノ酸(塩基では871番目)、AXは216番目のアミノ酸(塩基では646番目)、B3は352番目のアミノ酸(塩基では1054番目)にこうした違いが起きていた[81]。
基本的に型が同じなら抗原は同じもの(量が異なるのみ)なので亜型が違っても通常はその型の赤血球製剤で問題ない[82]し、反応する場合も低温でのみ反応する寒冷凝集素の場合は実害がないためそのまま輸血可能だが、まれに37℃反応性のその型の抗体(A型なら抗A1抗体、B型なら抗B抗体)を持っているばあいは「O型」の赤血球製剤(A抗原・B抗原を持たない)を使用する。いずれの場合も血漿・血小板剤はその型のもので問題ない[83]。
A型の亜型
- A1
- 普通のA型。A型の人のうち約80%を占める。(赤血球1個当たりの抗原数8.1×105〜11.7×105)[82]
- Aint
- A1よりも弱くA2よりも強い。
- A2
- 弱いA型。(赤血球1個当たりの抗原数2.4×105〜2.9×105)
- このあたりからO型に間違えられやすくなる[82]。
- 検査は抗Hレクチン、抗A1レクチン、被凝集価測定、転移酵素活性測定、唾液、血清中の型物質測定、
- そのほかA1に対する抗体を持つものものが時々いる[84]ため、A型血球との間接抗グロブリン試験などでも調べる。
- A3
- かなり弱いA型。(赤血球1個当たりの抗原数7000)
- オモテ試験で部分凝集となるのが特徴。
- 検査は抗Hレクチン、抗A1レクチン、被凝集価測定、転移酵素活性測定、唾液、血清中の型物質測定。
- その他A型とO型の血液キメラやモザイクとの鑑別のため、混合赤血球の分離も。
- これ以外にほとんどがA1に対する抗体を持つという性質を持つ[85]。
- Ax
- A3よりさらに弱いA型。(赤血球1個当たりの抗原数1400〜10300)
- AでありながらAに対する抗体を持ち、あるはずの転移酵素や型物質がない。
- 検査は抗Hレクチン、抗A1レクチン、吸着解離試験、転移酵素活性測定、唾液、血清中の型物質測定。
- Am
- Axよりさらに弱いA型。(赤血球1個当たりの抗原数1200)
- Aでありながらオモテ試験で凝集せずOと判定される。しかし転移酵素や型物質は存在する。
- 検査は抗Hレクチン、抗A1レクチン、吸着解離試験、転移酵素活性測定、唾液、血清中の型物質測定。
- Ael
- ものすごく弱いA型。(赤血球1個当たりの抗原数700)
- 「el」はelution(溶離・溶出)の略。吸着解離試験の検査以外ではA型と判断できない。
- Aend
- ものすごく弱いA型。特定の抗原が存在しないか、ごくわずかしか存在しない。
| 亜型 | 抗A血清との反応 | 抗B血清との反応 | 血清中の抗A | 血清中の抗B | 型物質 | A型転移酵素 | 適切な追加検査 |
|---|---|---|---|---|---|---|---|
| A1 | + | 0 | 0 | + | A、H | あり | なし |
| A2 | + | 0 | +/0 | + | A、H | あり | Hレクチンとの反応、A1レクチンとの反応、被凝集価測定、唾液・血清中の型物質測定、転移酵素活性測定、A血球との間接抗グロブリン試験 |
| A3 | mf(部分凝集) | 0 | 0 | + | A、H | あり | Hレクチンとの反応、A1レクチンとの反応、被凝集価測定、唾液・血清中の型物質測定、転移酵素活性測定、混合赤血球分離 |
| Ax | +/0 | 0 | + | + | H | なし | Hレクチンとの反応、A1レクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定、A血球との間接抗グロブリン試験、家系調査 |
| Am | 0 | 0 | 0 | + | A、H | あり | Hレクチンとの反応、A1レクチンとの反応、吸着解離試験、唾液・血清中の型物質測定、転移酵素活性測定、家系調査 |
| Ael | 0 | 0 | + | + | H | なし | Hレクチンとの反応、A1レクチンとの反応、吸着解離試験、唾液中の型物質測定、転移酵素活性測定、家系調査 |
基本的に血清中に抗Aがあると、血清を使った型物質測定はできない。
B型の亜型
B型はあまり研究が進んでいないが、A型同様のバリエーションがあると思われる。
- B1
- 普通のB型。
- Bint
- B1よりも弱くB2よりも強い。
- B2
- 弱いB型。
- B3
- かなり弱いB型。
- オモテ試験で部分凝集となるのが特徴。
- 検査は抗Hレクチン、被凝集価測定、転移酵素活性測定、唾液、血清中の型物質測定。
- そのほかキメラやモザイクとの鑑別のため、混合赤血球の分離も。
- Bx
- B3よりさらに弱いB型。
- BでありながらBに対する抗体を持ち、あるはずの転移酵素や型物質がない。
- 検査は抗Hレクチン、吸着解離試験、転移酵素活性測定、唾液、血清中の型物質測定。
- Bm
- Bxよりさらに弱いB型。
- Bでありながらオモテ試験で凝集せずOと判定される。しかし転移酵素や型物質は存在する。
- 検査は抗Hレクチン、吸着解離試験、転移酵素活性測定、唾液、血清中の型物質測定。
- Bel
- ものすごく弱いB型。特定の抗原が存在しないか、ごくわずかしか存在しない。
- 「el」は吸着解離試験を意味する。この検査以外ではB型と判断できない。
- Bend
- ものすごく弱いB型。特定の抗原が存在しないか、ごくわずかしか存在しない。
| 亜型 | 抗A血清との反応 | 抗B血清との反応 | 血清中の抗A | 血清中の抗B | 型物質 | B型転移酵素 | 適切な追加検査 |
|---|---|---|---|---|---|---|---|
| B | 0 | + | + | 0 | B、H | あり | なし |
| B3 | 0 | mf(部分凝集) | + | 0 | B、H | あり | Hレクチンとの反応、被凝集価測定、血清・唾液中の型物質測定、転移酵素活性測定、混合赤血球分離 |
| Bx | 0 | +/0 | + | + | H | なし | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
| Bm | 0 | 0 | + | 0 | B、H | あり | Hレクチンとの反応、吸着解離試験、血清・唾液中の型物質測定、転移酵素活性測定 |
| Bel | 0 | 0 | + | + | H | なし | Hレクチンとの反応、吸着解離試験、唾液中の型物質測定、転移酵素活性測定 |
基本的に血清中に抗Bがあると、血清を使った型物質測定はできない。
AB型の亜型
- 前述のA型亜型とB型亜型の組み合わせ
- 例えばA1B2の場合、Bの凝集力が弱いのでA型と誤認されやすくなる。両方凝集が弱い型だとO型との誤認もありうる[82]。
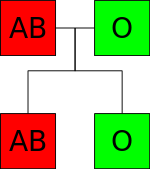
普通、AB型(遺伝子型はA/B)とO型(遺伝子型はO/O)の両親からはA型とB型のこどもが生まれるが、遺伝子型がcis-AB/OとO/Oの親からはAB型(シスAB)とO型のこどもしか生まれない。
シスAB型 (cisAB)- 普通、A型遺伝子とB型遺伝子が重なった際にAB型になる(例・A×B=AB)。しかし、シスAB型の人には、AB型遺伝子ともいえるものが存在し、配偶者がO型などでもAB型が生まれる事がある(例えばcisAB×Oの場合は全ての型が生まれる可能性がある)。ちなみに、普通のAB型はトランスAB型と呼ばれる。
- シスAB型の場合、普通のAB型に比べて抗原を作る量が少ないことが多く、A2B3(A・B双方の抗原が少ない)、A2B(B抗原は多いがA抗原は少ない)、A1B3(A抗原が多いがB抗原が少ない)の3種類が報告されており[82]、一番典型的なA2B3型の場合、A抗原・B抗原は弱いがH抗原は通常より高く、A1レクチンに反応しない。血清に弱い抗B抗体があり、時には抗A1抗体もあるが体温では反応しないなどの特徴を持つ[86]。
- 遺伝子を見るとA・B双方の抗原が少ないA2B3型の場合、A型遺伝子を基本に2か所(156番目と268番目)のアミノ酸が異なり、後者がB遺伝子の物と同じ配列になるのでAとBの酵素をつぎはぎにしたような酵素ができ、量は少ないがAとB双方の抗原が作られているのに対し、B抗原は多いがA抗原は少ない型の場合はB型遺伝子を基本にしているが235番目のアミノ酸がA型と同じ配置なのでB抗原が主に作られ、わずかだがA抗原も作られているといったような違いがある[87]。
| 遺伝子型 | 表現型 | 抗A血清との反応 | 抗B血清との反応 | 血清中の抗A | 血清中の抗B | 型物質 | 転移酵素 | 適切な追加検査 |
|---|---|---|---|---|---|---|---|---|
| AB/O | A2B3 | + | mf(部分凝集) | +/0 | + | A、(B)、H | なし | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定、混合赤血球分離 |
| AB/A | A1B3 | + | mf(部分凝集) | 0 | + | A、(B)、H | A | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定、混合赤血球分離 |
| AB/B | A2B | + | + | +/0 | 0 | A、B、H | B | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
O型の亜型(便宜上ボンベイ型・パラボンベイ型も解説)
- (本来の)O型の亜型
- 元々O型が「ABO遺伝子のうちH物質に糖をつける遺伝子が働かないもの」すべてを指すので遺伝子の配列がかなり違うものが見つかっており、通常の87番目のアミノ酸製造の塩基が1つ抜けているもののほかに、さらに後半部で置換がある亜型と、87番目のアミノ酸はA型やB型と同じだが後半のアミノ酸で塩基の置換が生じてアミノ酸が4つ(176・235・266・268番目)異なっている(厳密には176番目がB型、235・256番目がA型、268番目がどちらとも違う。)亜型も見つかっている[88]。
- いずれでも遺伝子を調べて分かる程度の違いでH物質が赤血球にそのままついているのには変わりがなく、輸血上の問題もない。
- ボンベイ型(Oh)
インドのボンベイ(現在のムンバイ)で発見されたことから、この名前がついている。- H物質を組み立てる際にフコースをつける工程があるが、ボンベイ型はこの酵素が作られない遺伝子のためH物質自体が完成せず、ここから先のA抗原とB抗原も作られないためA型やB型の遺伝子を持っていてもおもて試験ではO型と判定されてしまう。
- H物質を持たないため抗H抗体を自然抗体として持ち、うら試験で通常A・B型血球と対照用に使うO型血球[89]を凝集させる。
- この抗H抗体は体温で反応する[脚注 12]ので、ボンベイ型にO型を含むH抗原のある型の血液を輸血できず、同型(ボンベイ型)の赤血球製剤を輸血する[90]。
- 表記は「Oh」だが、O型血液を輸血できないことなどから厳密にはO型と全く別の血液型である[91]。
- パラボンベイ型(記号は下記参照)
- ボンベイ型と同様にH物質を組み立てる遺伝子の変異でH物質が完成しないが、こちらは赤血球にA抗原もしくはB抗原を弱くだが持つ型。(理由は#機構を参照)
- Ah型、Bh型(Row-IIとRow-IIIに分類[脚注 13])、AHm型[脚注 14]、BHm型、OHm型(Row-IIIに分類)が確認されている。
- Ah型、Bh型:通常のボンベイ型(Oh型)と同じ性質を持つが、AまたはB抗原が不完全で弱いながらも存在。
- AHm型、BHm型:血液上の抗原はAh型、Bh型と同様だが唾液は分泌性、OHm型はOh型(通常のボンベイ型)と同様だが唾液は分泌型。
- 輸血の問題はボンベイ型と基本的に同じだが、Row-IIIのパラボンベイ型で抗体が低温性のもの(抗HI)のみAhの場合はA型、Bhの場合はB型を使用してよい。血漿・血小板剤はその型(亜型の種類によってA・B・Oの選択肢がある)のもので問題ない[92]。
| H | Se | 表記 | 抗A血清との反応 | 抗B血清との反応 | 抗H血清との反応 | 唾液中のA型物質 | 唾液中のB型物質 | 唾液中のH型物質 | 血清中抗体 | 適切な追加検査 |
|---|---|---|---|---|---|---|---|---|---|---|
| 不活性 | 非分泌型 | Oh | 0 | 0 | 0 | 0 | 0 | 0 | 抗H | Hレクチンとの反応、吸着解離試験、唾液中の型物質測定、転移酵素活性測定 |
| 活性低下 | 非分泌型 | Ah | +/0 | 0 | +/0 | 0 | 0 | 0 | 抗H | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
| 活性低下 | 非分泌型 | Bh | 0 | +/0 | +/0 | 0 | 0 | 0 | 抗H | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
| 活性低下 | 分泌型 | Om | 0 | 0 | +/0 | 0 | 0 | + | 抗HI | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
| 活性低下 | 分泌型 | Am | +/0 | 0 | +/0 | + | 0 | + | 抗HI | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
| 活性低下 | 分泌型 | Bm | 0 | +/0 | +/0 | 0 | + | + | 抗HI | Hレクチンとの反応、被凝集価測定、唾液中の型物質測定、転移酵素活性測定 |
判定方法
試薬の抗A血清と抗B血清とを用いて、採取した赤血球と反応させて凝集の有無により判定する方法(おもて検査)で仮に判定される(抗H血清も使用することがある。抗H血清を使用するとボンベイ型の判定も出せる)。どちらかの血清で凝集が見られた場合はその血液型、どちらとも凝集が見られた場合はAB型、凝集が見られない場合はO型と判定される。これに加え、血液の血清を用いてA・B・O各自型の血球の凝集(O型血球は対照として用い、これが凝集する場合は判定を保留する。)を判定する方法(うら検査)で判定して結果が一致した場合に、血液型が確定される。
誕生時には、うら検査で判定するのに必要な血液型決定因子が不足しているので判定できず、おもて検査では、凝集が起きにくいタイプの場合や凝集の有無を間違って、誤って仮判定されるケースがある。そのため、成長してから正しい血液型が確定された場合に、ABO型の血液型が変わったかのように見える場合がある。なお、おもて検査とうら検査の判定が一致しなかった場合は再検査する。それでも一致しなかった場合は以下の可能性も考慮する。おもて検査とうら検査には優劣がないため、どちらかの判定を優先して血液型を決定するということはしない。
おもてとうら不一致時に考えられる可能性[93]。
- 血球側に問題がある場合の例
- 亜型(#ABO式血液型の亜型分類参照)
- 疾患による後天性の抗原異常(白血病やホジキン病などで抗原が弱まり、弱い亜型やO型と間違えやすくなる。)
- 獲得性B(A型が癌や細菌による感染症で発生したB抗原が赤血球につき、AB型に見える場合がある。)
- 血液キメラ・モザイク(複数の遺伝子や同じ遺伝子でも表現型が異なる血球が存在する。当然別型の血球ごとに反応が変わる。)
- 汎血球凝集反応(血球の表面が細菌やウイルスによって劣化し、T抗原系統の露出ですべての抗原血清で凝集する。#汎血球凝集の各レクチンに対する反応参照)
- 自己免疫性疾患による血球の抗体感作(自分の血液に反応する抗体が元からある)
- 異型輸血後(別人の血が入っているのでキメラ・モザイク同様に別型の血球で反応が異なる。)
- 血清側に問題がある場合の例
- 連銭形成(凝集とは別の赤血球が数珠上に重なった状態を凝集と誤認。)
- 低または無γグロブリン血症で抗体不足
不規則抗体の存在(ABO式と無関係の血液型で凝集)- 血清中の血液型物質の増加(癌などで見られる場合がある)
- 高力価の寒冷凝集素
- 新生児や老人における抗A抗B抗体の欠損または低下(上記参照)
- 新生児の胎盤通過性母親由来抗体(母親の抗体が新生児血清に混ざっている)
- 亜型血清中の抗体(亜型の一部には同型抗原に抗体を持つ場合がある)
血液ではなく、遺伝子から判定するという手法もあり、血清による判定に比べ、誤判定が生じにくいことが特徴である。
| 反応強度 | スコア | 特徴と外観 | 背景の色調 |
|---|---|---|---|
| 4+ | 12 | 一個の大きな凝集塊 | 透明 |
| 3+ | 10 | 数個の大きな凝集塊 | 透明 |
| 2+ | 8 | 中程度の凝集塊 | 透明 |
| 1+ | 5 | 小さな凝集塊 | 赤く濁る |
| w+ | 2 | ごくわずかな微小凝集 | 赤く濁る |
| 0 | 0 | 凝集も溶血もみられない | 赤く濁る |
| mf | 部分凝集 | 赤く濁る | |
| H(PH) | 完全溶血(部分溶血) | 赤く透明(濁る) |
また、亜型検査は、輸血検査の中でも血液型を確定するのに非常に重要である。
抗原側検査
- 抗Hレクチン
- H抗原のない、O型の亜型であるボンベイ型やパラボンベイ型には凝集せず、逆にその他の亜型には激しい凝集を起こさせる。またCad(+)血球にも凝集。
- 対象は全亜型。特にボンベイ型、パラボンベイ型
- 通常は
- O>A2B>B>A1>A1Bの順に凝集は強くなる。
抗A1レクチン(ドリコスレクチン[94])- A1抗原のない、A型の亜型であるA2型などには凝集しない。
汎血球凝集などでも例外的に凝集する。- 対象はAの亜型。
- ピーナッツレクチン
- 感染症などで細菌の酵素により血球の内在性抗原(T、Tk、Tnなど)が露出し、ピーナッツレクチンをはじめ全抗血清に凝集する。
- 対象は汎血球凝集を疑う場合。詳細は下記
- 吸着解離試験
- 血球に抗原が存在することを証明するために、既知の抗体をいったん患者血球に吸着させ、熱解離などによって再び解離する。そして既知の同型血球とまた反応するかを調べる。
- 対象は亜型のうちAm、Ax、Ael、Bm、Bx、Bel型。
- 再アセチル化
- 消化器系の感染症では細菌の酵素でN-アセチルガラクトサミンが脱アセチル化され、ガラクトースとなり抗B血清と反応するようになる。
無水酢酸で再アセチル化すれば通常の状態に戻る。- 対象は後天性B。
被凝集価測定- 抗原の強さを調べる。抗血清の希釈系列と患者血球を反応させ、どこまで凝集するかを調べる。
- 対照と比較して被凝集価が2管差以上、スコアが10以上あれば亜型。
- 対象はA2、A3、B3、パラボンベイ型
- 吸収試験
- 抗原の強さを調べる。被凝集価とは逆に患者血球の希釈系列と抗血清を反応させ、どこまで凝集するかを調べる。
- 吸収価が1:8未満ではA3、B3以外の亜型。
- 対象はA2、A3、B3、パラボンベイ型
- 混合赤血球の分離
- 対象は異型輸血、キメラ、モザイクを疑う場合。
- もともと部分凝集がみられた場合、「異型輸血」「亜型」「造血幹細胞移植後」「キメラ・モザイク」「白血病・ホジキン病などの疾患による抗原減弱」を疑う。
血清側検査
転移酵素活性測定- 血清中にあるはずの転移酵素の有無を調べる。
- 患者血清と試薬、O型血球を37度で反応させた後、O型血球が転移酵素の作用で別の型に変わったか被凝集価測定で確認する。
- 対象は全亜型。Ax、Ael、Bx、Belでは検出されない。
- 型物質測定(血清)
- 血清中にあるはずの型物質の有無を調べる。
- 抗血清の希釈系列と患者血清を混ぜ、さらに同型血球を反応させてどこまで凝集するかを調べる。
- もし型物質があればそれによって凝集が妨害を受ける。
- 型物質測定(唾液)
- 患者が分泌型(Lewis(a-b+))なら唾液中にも型物質があるので血清の代わりになる。
- Lewis(a-b+)は日本人の70パーセント。分泌型。
- Lewis(a+b-)は日本人の20パーセント。非分泌型、分泌型両方あり。
- Lewis(a-b-)は日本人の10パーセント。非分泌型。
- 対象は全亜型。Ax、Ael、Bx、BelやシスABでは検出されない。
不規則抗体検査- (不規則抗体はABO式血液型以外の血液型に対する抗体のことだが、便宜上ここに記す。)
- その他IgM系不規則抗体でもウラ試験で異常凝集がある。
- 対象はウラ試験で異常な凝集が認められた場合。
- 予備加温法によるウラ試験
- 37度の熱を加えてウラ試験の凝集が消えるか確認する。
- 対象は寒冷凝集を疑う場合。
- 生食置換法によるウラ試験
- 生理食塩水を加えてウラ試験の凝集が消えるか確認する。
- 対象は連銭形成を疑う場合。
解離試験・吸収試験
熱解離
赤血球沈渣(6回洗浄済)と生食を1容量:1容量混和。
56℃で10分加温後、900 - 1000G(3400rpm)で2分遠心し、上清を解離液とする。
| !目的 | 方法 | 解離液(性状) | 反応温度 | 反応時間 | 解離液(色) | 解離後血球の利用 | 試薬 |
|---|---|---|---|---|---|---|---|
| 解離 | 熱解離 | IgM | 50 - 56 | 5 - 10分 | 淡赤色 | 不可 | 生食 |
主にAm、Ax、Ael、Bm、Bx、Belなどの亜型に対し、抗原の存在を証明する吸着解離試験で実施。処理血球の検査はできない。
DT解離
赤血球沈渣(3回洗浄済)と生食、DT液を1容量:1容量:2容量混和。
37℃で5分反応後、900 - 1000G(3400rpm)で5分遠心し、上清を解離液とする。
ジキトニン解離
赤血球沈渣(6回洗浄済)と生食を1:9混和。
0.5%ジキトニン液0.5mlを加え1分転倒混和し、900 - 1000G(3400rpm)で5分遠心。赤血球残渣が白くなるまで5回以上洗浄(2分遠心)。
赤血球残渣に0.1Mグリシン塩酸緩衝液2.0mlを混和し1分転倒混和。
900 - 1000Gで5分遠心し、解離液に0.8Mリン酸緩衝液0.2mlを加え、さらに2分遠心し、上清を解離液とする。
目的方法||解離液(性状)反応温度反応時間解離液(色)解離後血球の利用試薬
| 解離 | エーテル解離 | IgG | 37 | 30 - 40分 | 暗赤色 | 不可 | エチルエーテル |
| 解離 | DT解離 | IgG | 37 | 5分 | 暗赤色 | 不可 | ジクロロメタン・ジクロロプロパン |
| 解離 | ジキトニン酸解離 | IgG | 室温 | 1分 | 無色 | 不可 | ジキトニン液、グリシン塩酸、リン酸緩衝液 |
主に直接クームス陽性血球から、抗体が含まれる解離液を入手するために実施。処理血球の検査はできない。酸解離では酵素法で検出するはずの抗体は検出できない。
グリシン塩酸/EDTA解離
0.1Mグリシン塩酸緩衝液とEDTA溶液を20容量:5容量混和。
赤血球沈渣(6回洗浄済)とグリシン塩酸/EDTA液を10容量:20容量混和。
室温で2 - 3分反応後、1Mトリス/NaClを1容量混和。
900 - 1000G(3400rpm)で2分遠心し、上清を解離液とする。処理血球は生食で4回洗浄し使用。
| 目的 | 方法 | 解離液(性状) | 反応温度 | 反応時間 | 解離液(色) | 解離後血球の利用 | 試薬 |
|---|---|---|---|---|---|---|---|
| 解離 | グリシン塩酸・EDTA解離 | IgG | 室温 | 2 - 3分 | 無色 | 可 | グリシン塩酸・EDTA、1Mトリス/NaCl |
抗体解離液が得られ、処理血球の検査も可能。
クロロキン解離法
赤血球沈渣(6回洗浄済)と20%クロロキン2リン酸を1:4混和。
室温で30分反応後、3回以上洗浄し血液型検査に用いる。
感作抗体だけでなく、Bg抗原を破壊できる。
| 目的 | 方法 | 解離液(性状) | 反応温度 | 反応時間 | 解離液(色) | 解離後血球の利用 | 試薬 |
|---|---|---|---|---|---|---|---|
| 解離 | クロロキン解離 | 解離液なし | 37 | 30分 | 解離液なし | 可 | クロロキン2リン酸 |
血液型検査を目的とした抗体解離法(抗体解離液は得られない)
ZZAP処理
赤血球沈渣とZZAPを1:2で混和し37℃30分間反応。生食で3回洗浄。
| 目的 | 方法 | 解離液(性状) | 反応温度 | 反応時間 | 解離液(色) | 解離後血球の利用 | 試薬 |
|---|---|---|---|---|---|---|---|
| 解離 | ZZAP処理 | 解離液なし | 室温 | 30分 | 解離液なし | 可 | DTT・フィシン |
自己抗体があると疑われた際、行われる自己抗体吸着のための抗体解離法
PEG吸収
自己血球:自己血清:PEG = 1:1:2で37℃15分間反応させ、900 - 1000G(3400rpm)で5分遠心し、上清を吸収液とする。
| 目的 | 方法 | 解離液(性状) | 反応温度 | 反応時間 | 解離液(色) | 解離後血球の利用 | 試薬 |
|---|---|---|---|---|---|---|---|
| 吸収 | PEG吸収 | 解離液なし | 37 | 解離液なし | 解離液なし | 解離液なし | ポリエチレングリコール |
自己抗体があると疑われた際に直接、自己抗体を吸着させる方法。ZZAPを使うより簡単だが、自己抗体以外の同種抗体を吸着させてしまうこともある。
また、寒冷凝集の場合は試薬は要らず、全血で4℃に1時間保冷すれば自己吸着できる。
汎血球凝集の各レクチンに対する反応
感染症などによってTなどの内在性抗原が露出した血球は、どんな血清・血漿とも反応するようになる。これを汎血球凝集といい、血漿成分を含む製剤の輸血は避ける。
| 種類 | T | Tk | Th | Tx | Tn | Cad | HEMPAS | 正常 |
|---|---|---|---|---|---|---|---|---|
| 原因 | 感染症 | 感染症 | 感染症・血液疾患 | 感染症 | 血液疾患 | 遺伝 | 遺伝 | |
| 発現期間 | 一過性 | 一過性 | 一過性・長期 | 一過性 | 長期 | 永久 | 永久 | |
| Arachis hypogaea(ラッカセイ) | + | +(酵素処理で強化) | +(酵素処理で減弱) | + | 0 | 0 | 0 | 0 |
| Salvia sclarea | 0 | 0 | 0 | 0 | + | 0 | 0 | 0 |
| Salvia horminum | 0 | 0 | 0 | 0 | + | + | 0 | 0 |
| Glycine soja(ツルマメ) | + | 0 | 0 | 0 | + | +/0 | + | 0 |
| Vicia cretica | + | 0 | + | 0 | 0 | 0 | 0 | 0 |
| Griffonia simplifolia | 0 | + | 0 | 0 | 0 | 0 | 0 | 0 |
| Dolichos biflorus | 0 | 0 | 0 | 0 | + | + | 0 | 0 |
| ポリブレン | 0 | + | + | + | +/0 | + | + | + |
ABO型における親子の理論的な血液型の組み合わせ

A型(AO)とB型(BO)による組み合わせ
※あくまでも、メンデルの法則に基づいた、単純化した理論による血液型および確率である。現実には亜型等による例外が存在する。(例・シスAB型とO型によるAB型やO型の子供など)
ABO式血液型は、人の第9番染色体に存在する複対立遺伝子によって決定する。通常、存在する遺伝子の遺伝子型はA、B、Oの3種類であって、AとBとはOに対して優性に遺伝し、AとBとの間には優性劣性の差異は存在しない。すなわち、2本の第9番染色体のうち少なくとも一方にA遺伝子が存在しいずれにもB遺伝子が存在しなければ表現型はA型、少なくとも一方にB遺伝子が存在しいずれにもA遺伝子が存在しなければ表現型はB型、A遺伝子・B遺伝子の双方が存在すれば表現型はAB型、2本の染色体の双方にO遺伝子が含まれる場合は劣性遺伝するO型が表現型となる。
下の表のように、表現型がA型とB型の場合は複数の遺伝子型が存在する。
| A型 | B型 | O型 | AB型 | ||||
|---|---|---|---|---|---|---|---|
| AA | AO | BB | BO | ||||
| A型 | AA | A型 (AA) 100% | A型 100% (AA 50%、AO 50%) | AB型 100% | A型 (AO) 50%、 AB型 50% | A型 (AO) 100% | A型 (AA) 50%、 AB型 50% |
| AO | A型 100% (AA 50%、AO 50%) | A型 75% (AA 25%、AO 50%)、 O型 25% | B型 (BO) 50%、 AB型 50% | A型 (AO) 25%、 O型 25%、 B型 (BO) 25%、 AB型 25% | A型 (AO) 50%、 O型 50% | A型 50% (AA 25%、AO 25%)、 B型 (BO) 25%、 AB型 25% | |
| B型 | BB | AB型 100% | B型 (BO) 50%、 AB型 50% | B型 (BB) 100% | B型 100% (BB 50%、BO 50%) | B型 (BO) 100% | B型 (BB) 50%、 AB型 50% |
| BO | A型 (AO) 50%、 AB型 50% | A型 (AO) 25%、 O型 25%、 B型 (BO) 25%、 AB型 25% | B型 100% (BB 50%、BO 50%) | B型 75% (BB 25%、BO 50%)、 O型 25% | B型 (BO) 50%、 O型 50% | B型 50% (BB 25%、BO 25%)、 A型 (AO) 25%、 AB型 25% | |
| O型 | A型 (AO) 100% | A型 (AO) 50%、 O型 50% | B型 (BO) 100% | B型 (BO) 50%、 O型 50% | O型 100% | A型 (AO) 50%、 B型 (BO) 50% | |
| AB型 | A型 (AA) 50%、 AB型 50% | A型 50% (AA 25%、AO 25%)、 B型 (BO) 25%、 AB型 25% | B型 (BB) 50%、 AB型 50% | B型 50% (BB 25%、BO 25%)、 A型 (AO) 25%、 AB型 25% | A型 (AO) 50%、 B型 (BO) 50% | A型 (AA) 25%、 B型 (BB) 25%、 AB型 50% | |
ABO式血液型と性格
科学的には血液型と性格に関係があるとはされておらず、現時点で知られている血液型性格分類はいずれも正しいとは認められていない[95]。だが1970年代から2000年代前半にかけて、多くのテレビや書籍が根拠なく分類を広めたため、いまだに血液型と性格の関連性を信じている人もいる[96]。血液型性格分類が広まっているのは、日本とその影響を受けた韓国、台湾といった一部地域だけであり、それ以外の地域では性格と血液型を関係づける習慣がなく、日本の血液型性格分類は奇妙に思われている[97]。そもそも血液型への関心自体が薄く、自分の血液型を覚えていない人も多い(輸血が必要な時などは、その場で血液型検査が行われる)[98]。
血液型によって人の性格を判断し、相手を不快や不安な状態にさせる言動はブラッドタイプ・ハラスメント(通称ブラハラ)と呼ばれ、近年になり社会問題として取り上げられるようになった[99]。採用試験の応募用紙に血液型の記入欄があったため、改善するよう労働局から指導された企業もある[100]。厚生労働省は「血液型は職務能力や適性とは全く関係ない」と呼びかけている[100][101]。
一方で、免疫系の観点から血液型性格分類を支持する研究[102][103]や、日本の健常人において、ABO式血液型の遺伝子型と性格特性には有意な関連が認められるとする研究もある[104]。
ABO式血液型と体質
血液型と病気の関連性については1980年代には持てはやされていたが、ヒトゲノム計画が終りつつあった2000年に、科学雑誌『Nature』にて総説が掲載され、その内容は「胃腸管に関するいくつかの形質に弱い相関が確認できるが、血液型と疾患の相関については再現性よく示されたものは無い」というものであった[105]。その後の研究では、健康面(ストレス抵抗性や病気のリスク)へ影響があるという報告は存在している[106]。
ABO式以外の血液型では特定の血液型のみある病気にかからないというものがいくつか見つかっている(P式血液型:p型とPk型のヒトパルボウイルスB19耐性、ダフィー式血液型:Fy(a-b-)型の三日熱マラリア耐性など[107]。)が、ABO式では完全耐性の病気は見つかっておらず、下記のように相対値で数割ほど多い少ない程度の「あえて言えば」の範囲である。
- マラリア:マラリアそのものの感染率は変わらないが、熱帯熱マラリアによって起きる脳性マラリアに移行するしやすさがA型がO型の1.3倍。(マラリアに寄生された赤血球が脳の血管に詰まるのが原因だがA型のみ毛細血管に若干くっつきやすい。)
- 胃(十二指腸)潰瘍:ピロリ菌の感染しやすさがO型のみ25%ほど高い(ピロリ菌はルイス抗原のb抗原につきやすいので、Aルイスb抗原やBルイスb抗原に変化しているA型やB型より、これがこのまま残るO型の胃壁につきやすい。)
- ノロウィルス:一部の株に限るが、H物質につくのでO型の感染率が若干高い。
(以上3例山本(2015) p.180-188より)
免疫機構以外の病気の成りやすさで確認されているものでは、O型は血液凝固に必要なフォン・ウィルブランド因子の濃度が他の型より25%ほど低く、このため他の型よりわずかに血液が凝固しにくく、血栓が起きにくい(エコノミークラス症候群発生率がO型を基準とすると他の型は50%ほど多い)[108]。
ABO式血液型の変化
後天的な変化
ルイス式血液型では出生後に血液型は変化するがABO式では稀である。
白血病の治療などで造血幹細胞移植(骨髄移植)を行った場合には、移植したドナーの造血幹細胞によって血液を造り出すようになるため、原則としてドナーのABO式血液型に変わる。- 骨髄性白血病などで、特定の抗原糖の産出が停止し、血液型が変わることがある。
- 細菌感染症で、細菌が出すジアセチラーゼにより抗原糖が変質し、血液型が変わることがある。
ただし、上述の病気や細菌感染症で変わることは非常に稀である。現在の知見では病気やその治療以外の原因で血液型が変化することは基本的にありえないので、病気や治療などの原因がないにも関わらず献血等で血液検査を行ったときに血液型が異なっていた場合は、本人や親の単純な思い込みや新生児での血液検査が間違っていたと考えた方が良い。
輸血用ABO抗原の変更
(分泌型なら)唾液中からも血液型物質が検出できるのだが、すぐに調べればわかるのに数時間ほど放置してから調べると検出不能になる事例がかなり初期から知られており、さらに採集後すぐに加熱処理すれば長時間置いても検出できるため京城帝国大学の佐藤武雄 (法医学者)らはこれを研究した結果「口腔内の細菌が唾液中の血液型物質を分解する」ということを発見した。これはすべての型を分解するものだったが1950年代に井関尚栄らが選択的にA・B・Oの各型質を選択的に分解する酵素を発見し、例えばB型血球にB分解酵素を使うとO型血球になることが分かった(O型血球はフコースを失い、ガラクトースだけの志賀赤痢菌などに見られる異性人血球抗体を持つ血球になった。)[109]。
上記の時点では輸血に使える代物ではなかったが、2007年4月にA型、B型、AB型の赤血球をO型に変えることのできる酵素の開発に米国のハーバード大学などの国際研究チームが成功した。O型の血液はボンベイ型を除く全ての血液型の人に輸血が可能であるため、この技術が確立すれば、輸血の際に血液型を考慮する必要がほとんどなくなることとなる[110]。
脚注
^ 出典元では「ランドスタイナー」表記
^ 現在の観点から見ると不自然な親子関係があるが、参考にした『血液型の話』14Pの表のままで掲載。
^ あくまで二対対立因子説による仮説である点に注意
^ 前述のフォン・デュンゲルンおよびヒルシュフェルトの初めての調査ではAB型の親とO型の子の組み合わせがあるが、以後の他の学者たちによる調査で確認できなかった。
^ この違いはAB型に現れるはずで、ベルンシュタインの説では「AB型血球にはR凝集原がないので凝集素ρには反応しない」はずだが、古畑らの説は「O型血球に反応する凝集素があればAB型でも反応する」となる。
^ よってオーストラリア・南北アメリカ大陸などは先住民のデータから推測。
^ 比率でわかるように後述の「A型遺伝子やB型遺伝子の多い地域」でも大半はA型遺伝子やB型遺伝子は過半数に達しておらず、O型の方が多数派の場合も多い。
^ 参考にした『血液型の話』原文は「アメリカ・インディアン」表記
^ 古畑(1962) p.213
^ 日本やシベリア先住民ではやや低いが、北海道などのアイヌはB型が比較的多い。
^ A型遺伝子出現率は九州東部・四国南西部では29%以上だが、青森県の北部は24%以下になるなど(保志(1968) p.324図4「日本におけるA型遺伝子出現率の地域差」(古畑より改変)。)。
^ 抗H抗体自体はA型・AB型で一番多いA1型やA1B型の血清にも存在するが、こちらは体温で反応しないため輸血で問題にされることはほとんどない。
^ なお、Row-Iは通常のボンベイ型が分類される。
^ 注:Hを小文字で上に書く書き方もある、以下同じ
出典
^ 古畑(1962) p.9
- ^ abc 小川聡 総編集 『内科学書』第7版-vol6、中山書店、2009年、p49
^ 山本(2015) p.2-6
^ Dr. Karl Landsteiner: "Ueber Agglutinationserscheinungen normalen menschlichen Blutes", Wiener klinische Wochenschrift, 14 Jg., Nr.46 (14. November 1901), S.1132-1134.
^ Dr. Alfred v. Decastello und Dr. Adriano Sturli: "Ueber die Isoagglutinine im Serum gesunder und kranker Menschen", Münchener medicinische Wochenschrift, 49 Jg., No.26 (1. Juli 1902), S.1090-1095.
^ Prof. E. v. Dungern und Dr. L. Hirschfeld: "Ueber Vererbung gruppenspezifischer Strukturen des Blutes, II", Zeitschrift für Immunitätsforschung und experimentelle Therapie, 1 Teil, Bd.6, H.1 (22. Juni 1910), S.284-292.
^ yasuoka (2007年11月1日). “O型はゼロ型なのか | yasuokaの日記 | スラド”. yasuokaの日記. 2017年11月1日閲覧。
^ Klinische Wochenschrift, 7 Jg., Nr.35 (26. August 1928), S.1671.
^ 古畑(1962) p.14 I・2表
^ 古畑(1962) p.16 I・3表
^ 古畑(1962) p.16 I・4表
^ 古畑(1962) p.21
^ 古畑(1962) P.14-29「ABO式血液型の遺伝」
^ 古畑(1962) p.29-32
^ 山本(2015) p.44-51
^ 山本(2015) p.63-72
^ 山本(2015) p.97-101
^ “大阪大病院、輸血ミスで患者死亡 新鮮凍結血漿”. 47NEWS (2010年4月8日). 2012年8月15日閲覧。
^ ただしニワトリは元々ABO式の遺伝子を持ってない(山本(2015) p.163表ABO血液型が分かる科学』P163表)。
^ 山本(2015) p.124-125
^ ヨーロッパはM遺伝子とN遺伝子が半々ぐらいだが、北米先住民はM型、オーストラリア先住民はN型に偏っている。
保志(1968) p.326図1「M型遺伝子頻度の地理的分布」
^ 保志(1968) p.324-325
^ 保志(1968) p.324図1~3「O・A・B型の出現率の地域差」(ムーラントより改変)。
^ סוגי דם
^ [Blood-group systems ABO and RH in the Kenyan population].
^ BLUTGRUPPEN & RHESUSFAKTOR
^ 台人O型血佔4成 AB型僅6%
^ Türkiye'de en yaygın kan grubu A Rh pozitf
^ blodtype
^ Trend of blood groups and Rh Factor in the twin cities of Rawalpindi and Islamabad
^ STUDY OF ABO AND RH-D BLOOD GROUPS AMONG THE COMMON PEOPLE OF CAPITAL CITY OF BANGLADESH
^ Importance of Blood Typing
^ What's Your Type?
^ Rotes Kreuz: Blutgruppen
^ FAQ | Croix-Rouge luxembourgeoise
^ SPR Veripalvelu - Veriryhmät ja niiden periytyminen
^ HEMOAM - Fundação Hospitalar de Hematologia e Hemoterapia do Amazonas
^ Blood Types | National Blood Transfusion Service
^ Give Blood - Blood around the World
- ^ abcdAVIS Comunale Forlì - I gruppi sanguigni
^ Preguntas Frecuentes
^ CIA World Factbook
^ Blood Types - What Are They?, Australian Red Cross
^ “Austrian Red Cross - Blood Donor Information”. Old.roteskreuz.at (2006年3月21日). 2010年11月19日閲覧。
^ “Rode Kruis Wielsbeke - Blood Donor information material”. Rodekruiswielsbeke.be. 2010年11月19日閲覧。
^ Tipos Sanguíneos
^ Canadian Blood Services - Société canadienne du sang. “Types & Rh System, Canadian Blood Services”. 2010年11月19日閲覧。
- ^ abhttp://bloodtypes.jigsy.com/East_Asia-bloodtypes
^ Czech Red Cross. “Podíl krevních skupin v populaci České republiky”. 2011年3月18日閲覧。
^ Frequency of major blood groups in the Danish population.[リンク切れ]
^ http://www.verekeskus.ee/?op=body&id=27
^ “Suomalaisten veriryhmäjakauma” (Finnish) (2009年8月21日). 2010年11月19日閲覧。
^ “Les groupes sanguins (système ABO)” (French). Centre Hospitalier Princesse GRACE - Monaco. C.H.P.G. MONACO (2005年). 2008年7月15日閲覧。
^ “Blóðflokkar” (Icelandic). .landspitali.is. 2010年11月19日閲覧。
^ “ABO and Rh (D) group distribution and gene frequency; the first multicentric study in India”. 2016年4月25日閲覧。
^ “Irish Blood Transfusion Service - Irish Blood Group Type Frequency Distribution”. Irish Blood Transfusion Service. 2009年11月7日閲覧。
^ “The national rescue service in Israel” (Hebrew). Mdais.org. 2010年11月26日時点のオリジナルよりアーカイブ。2010年11月19日閲覧。
^ “血液の知識”. bloodbook.com. 2017年7月31日閲覧。
^ “Voorraad Erytrocytenconcentraten Bij Sanquin” (Dutch). 2009年3月27日閲覧。
^ “What are Blood Groups?”. NZ Blood. 2010年11月19日閲覧。
^ “Hva er en blodtype?” [What is a blood type?] (Norwegian). 2014年9月23日閲覧。
^ “Philippine Red Cross”. 2014年7月15日閲覧。
^ “Regionalne Centrum Krwiodawstwa i Krwiolecznictwa we Wroclawiu” (Polish). Rckik.wroclaw.pl (2010年9月2日). 2010年11月19日閲覧。
^ “Portuguese Blood Institute” (Portuguese). 2016年4月25日閲覧。 (assuming Rh and AB antigens are independent)
^ “Frequency of ABO blood groups in the eastern region of Saudi Arabia”. Cat.inist.fr. 2010年11月19日閲覧。
^ “South African National Blood Service - What's Your Type?”. Sanbs.org.za. 2010年11月19日閲覧。
^ “Federación Nacional de Donantes de Sangre/La sangre/Grupos”. Donantesdesangre.net. 2010年11月19日閲覧。
^ “Frequency of major blood groups in the Swedish population”. Geblod.nu (2007年10月2日). 2010年11月19日閲覧。
^ “Turkey Blood Group Site”. Kangrubu.com. 2010年11月19日閲覧。
^ “Blood Group Basics”. Blood.co.uk. 2014年5月3日閲覧。
^ Blood Types in the U.S - ウェイバックマシン(2011年7月19日アーカイブ分)
^ “Racial and ethnic distribution of ABO blood types”. Bloodbook.com. 2010年8月1日閲覧。
^ 古畑(1962) p.181 VIII・1表
^ 古畑(1962) p.183 VIII・2表
^ 古畑(1962) p.178-184
^ 古畑(1962) p.178-180
^ 古畑(1962) p.193
^ 古畑(1962) p.34
^ 古畑(1962) p.P193-194「5 アメリカ・インディアンの研究」
^ 古畑(1962) p.P194-195
^ 山本(2015) p.78-84
- ^ abcde松尾(2009) p.116
^ 北村(2006) p.344-345
^ 山本(2015) p.80-81
^ 山本(2015) p.80-81
^ 北村(2006) p.344・346
^ 山本(2015) p.84-89
^ 山本(2015) p.89-92
^ 北村(2006) p.345
^ 北村(2006) p.344-345
^ 山本(2015) p.103
^ 北村(2006) p.344-345
^ 北村(2006) p.345-346
^ 山本(2015) p.78-79
^
- 縄田健悟(2014)「血液型と性格の無関連性 ―日本と米国の大規模社会調査を用いた実証的論拠―」『心理学研究』85(2), 148-156. - 日米約1万人のデータを調べたが有意差は出なかった。
- 上村晃弘、サトウタツヤ(2006)「疑似性格理論としての血液型性格関連説の多様性」『パーソナリティ研究』15(1), 33-47.
- 広島修道大学人文学部助教授 中西大輔のウェブサイト「血液型性格判断をやめよう」 - 血液型性格判断の持つ問題点や差別性が心理学者の立場から詳説されている。
- 大阪大学大学院 生命機能研究科 認知脳科学研究室「血液型と性格は関係があるか?」
大村政男(1990, 1998新訂, 2012新編)『血液型と性格』福村出版- 村上宣寛(2005)『「心理テスト」はウソでした。受けたみんなが馬鹿を見た』日経BP社
- WU Kunher, LINDSTED Kristian D., LEE Jerry W. (2005). Blood type and the five factors of personality in Asia. - 台湾の論文。この調査では、血液型と外向性の関連性を否定する結果が出た。
- 山崎賢治・坂元章(1992) 「血液型ステレオタイプによる自己成就現象〜全国調査の時系列分析〜」『日本社会心理学会第33回大会発表論文集』 - 血液型性格関連説が社会的に広まり始めた数年後の1978年を起点に1988年まで、日本人延べ32,347人の自己評価による性格の経年変化を調べた。血液型性格分類を信じ、自分の性格をそれに合わせて振る舞っている人が有意に多いことを示した。
- Sakamoto, A., & Yamazaki, K. (2004). Blood-typical personality stereotypes and self-fulfilling prophecy: A natural experiment with time-series data of 1978–1988. Progress in Asian Social Psychology, 4, 239–262. - 上記の山崎賢治・坂元章(1992)と同様の内容。
- 武藤浩二・長島雅浩他(2012)「教員養成課程における科学リテラシー構築に向けた疑似科学の実証的批判的研究」『2011年度科研費研究成果報告書』 - 山崎賢治・坂元章(1992)は1978年から1988年までの11年間に毎年約3,000人(延べ32,347人)を解析したものであるが、このデータを2000年代にまで拡張して解析しても、同様の結果がでることが判明した(詳細な人数・年数は報告書には未掲載)。
- 山岡重行(2009)「血液型性格判断の差別性と虚妄性(自主企画(2))」『日本パーソナリティ心理学会大会発表論文集』 - 1999年から2009年までの6600人を調べたところ、血液型性格分類に対するステレオタイプを持つ被験者に限り多くの項目で有意差が出た。
^ MARI YAMAGUCHI (2005年5月6日). “Myth about Japan blood types under attack”. The Canadian Press. 2013年2月15日時点のオリジナルよりアーカイブ。2014年4月5日閲覧。
^ 海外記事を紹介するnewSphereの記事(血液型に対する海外メディアの反応)では「海外では血液型と性格を結びつけることはないためか、日本のこうした慣習は新鮮なようだ」としている。しばしば海外メディアは日本の奇妙(Weird)な習慣として血液型性格診断を紹介している(米NYタイムズ記事、英BBC記事)
^ BBC NEWS Japan and blood types: Does it determine personality?
^ “「血液型を扱う番組」に対する要望”. BPO(放送倫理・番組向上機構)青少年委員会. 2014年4月5日閲覧。
- ^ ab縄田健悟(2014)「血液型と性格の無関連性 ―日本と米国の大規模社会調査を用いた実証的論拠―」『心理学研究』85(2), 148-156.
“科学 血液型と性格「関連なし」…日米1万人超を調査”. 読売新聞(YOMIURI ONLONE). http://www.yomiuri.co.jp/kyoiku/news/20140722-OYT8T50051.html 2014年8月17日閲覧。
“B型はマイペース?…研究者「関係ありません」”. YOMIURI ONLINE KODOMO. http://www.yomiuri.co.jp/kodomo/newspaper/compare/20140723-OYT8T50076.html?from=yartcl_popin 2014年8月17日閲覧。
^ 厚生労働省の「血液型は職務能力や適性とは全く関係ない」という発表内容は熊本労働局のサイトで確認できる。2015年10月12日閲覧
ただし、全国の労働局を総括する厚生労働省本省のサイトでは同様の発表内容は確認できない。2015年10月19日閲覧
^ 藤田紘一郎(2006)『パラサイト式血液型診断』新潮選書
^ 藤田紘一郎(2010)『血液型の科学』祥伝社新書
^ 武田真貴子「本当だった!? 血液型による性格の違い」ケアネット、2015年6月2日
^ Risch NJ. (2000). Searching for genetic determinants in the new millennium. Nature, 405(6788), 847-856.
^
A Study on the effects of one's blood type on emotional character and antistress of adults, Journal of the Korea Academia-Industrial cooperation Society, 12(6), 2554-2560. - 2011年にソウルベンチャー情報大学院大学の金らが4000人以上の成人の脳波を測定した。2006年9月から2009年12月までに韓国精神科学研究所に脳波測定を依頼した成人(20歳〜59歳の4636人)のデータを分析した結果、O型は他の血液型に比べて注意指数や抗ストレス指数が有意に高かった。- 永田宏(2013)『血液型で分かる なりやすい病気 なりにくい病気 がん、胃潰瘍、脳梗塞から感染症まで』講談社ブルーバックス
- Brian M. Wolpin et al. (2009). ABO Blood Group and the Risk of Pancreatic Cancer. Journal of the National Cancer Institute, 101(6), 424-431. - 膵臓がんはB型はO型に比べると1.72倍リスクが高いなど。
^ 松尾友香、『最新 血液型の基本と仕組み』株式会社秀和システム、2009年、ISBN 978-4-7980-2422-6、P195-208。
^ 山本(2015) p.190-191
^ 古畑(1962) p.122-127。
^ Geoff Daniels; Stephen G Withers (2007年). “Towards universal red blood cells”. Nature Biotechnology 25: 427-428. http://www.nature.com/nbt/journal/v25/n4/full/nbt0407-427.html.
参考文献
- 古畑種基「血液型の話」、岩波書店、1962年。
- 保志宏、今尾恵介(監修)「機能から見た変異「血液成分の個人差」」、『原色現代科学大事典6-人間』、株式会社学習研究社、1968年、 323-327P。
北村聖、編 『看護のための最新医学講座』 (第2版版) 株式会社中山書店、2006年。ISBN 4-521-62511-8。
松尾友香 『最新 血液型の基本と仕組み』 株式会社秀和システム、2009年。ISBN 978-4-7980-2422-6。
- 山本文一郎 『ABO血液型が分かる科学』 株式会社岩波書店、2015年。ISBN 978-4-00-500811-7。
関連項目
血液型
ヒト白血球型抗原(HLA型、白血球の血液型)- Rh因子
- 認定輸血検査技師
- 血液型性格分類
- 血液型占い
外部リンク
【日本赤十字社】寄付・献血・ボランティア|血液の基礎知識 - 日本赤十字社サイト内


